
Как называется оплодотворение цветковых растений. Оплодотворение цветка
Двойное оплодотворение , половой процесс у покрытосеменных растений, при котором оплодотворяются как яйцеклетка, так и центральная клетка зародышевого мешка. Двойное оплодотворение открыл русский учёный С. Г. Навашин в 1898 на 2 видах растений - лилии (Lilium martagon ) и рябчике (Fritillaria orientalis ). В двойном оплодотворении участвуют оба спермия, привносимые в зародышевый мешок пыльцевой трубкой; ядро одного спермия сливается с ядром яйцеклетки, ядро второго - с полярными ядрами или со вторичным ядром зародышевого мешка. Из оплодотворённой яйцеклетки развивается зародыш, из центральной клетки - эндосперм. В зародышевых мешках с трёхклеточным яйцевым аппаратом содержимое пыльцевой трубки обычно изливается в одну из синергид, которая при этом разрушается (в ней видны остатки ядра синергиды и вегетативного ядра пыльцевой трубки); вторая синергида впоследствии отмирает. Далее оба спермия вместе с измененной цитоплазмой пыльцевой трубки перемещаются в щелевидный промежуток между яйцеклеткой и центральной клеткой. Затем спермии разобщаются: один из них проникает в яйцеклетку и вступает в контакт с её ядром, другой - проникает в центральную клетку, где контактирует со вторичным ядром или с одним, а иногда и с обоими полярными ядрами. Спермии теряют свою цитоплазму ещё в пыльцевой трубке или при проникновении в зародышевый мешок; иногда спермии в виде неизмененных клеток наблюдаются и в зародышевом мешке.
При двойном оплодотворении ядра зародышевого мешка находятся в интерфазе и обычно значительно крупнее ядер спермиев, форма и состояние которых могут вырьировать. У скерды и некоторых др. сложноцветных ядра спермиев имеют вид двойной скрученной или извитой хроматиновой нити, у многих растений они удлинённые, иногда извитые, более или менее хроматизированные, не имеющие ядрышек; обычно спермии представляют собой округлые интерфазные ядра с ядрышками, иногда не отличающиеся по структуре от женских ядер.
По характеру объединения мужских и женских ядер предложено (Е. Н. Герасимова-Навашина) различать два типа двойного оплодотворения: премитотическое - ядро спермия погружается в женское ядро, хромосомы его деспирализуются; объединение хромосомных наборов обоих ядер происходит в интерфазе (в зиготе); постмитотическое - мужское и женское ядра, сохраняя свои оболочки, вступают в профазу, в конце которой начинается их объединение; интерфазные ядра, содержащие хромосомные наборы обоих ядер, образуются лишь после первого митотического деления зиготы. При двойном оплодотворении в яйцеклетке сливаются 2 гаплоидных ядра, поэтому ядро зиготы диплоидно. Число хромосом в ядрах эндосперма зависит от числа полярных ядер в центральной клетке и от их плоидности; у большинства покрытосеменных 2 гаплоидных полярных ядра и эндосперм у них триплоиден. Следствие двойного оплодотворения - ксении - проявление доминантных признаков эндосперма отцовского растения в эндосперме гибридных семян. Если в зародышевый мешок проникает несколько пыльцевых трубок, спермий первой из них участвуют в двойном оплодотворении, спермий остальных - дегенерируют. Случаи диспермии, т. е. оплодотворения яйцеклетки двумя спермиями, очень редки.
Оплодотворению у покрытосеменных предшествует микро- и мегаспорогенез , а также опыление .
Микроспорогенез протекает в пыльниках тычинок. При этом диплоидные клетки образовательной ткани пыльника в результате мейоза превращаются в 4 гаплоидные микроспоры. Через некоторое время микроспора приступает к митотическому делению и преобразуется в мужской гаметофит – пыльцевое зерно .
Пыльцевое зерно снаружи покрыто двумя оболочками:экзиной и интиной . Экзина – верхняя оболочка более толстая и пропитана спороленнином – жироподобным веществом. Это позволяет пыльце выдерживать существенные температурные и химические воздействия. В экзине находятся проростковые поры, до опыления закрытые «пробочками». Интина содержит целлюлозу и эластична. В пыльцевом зерне имеются две клетки: вегетативная и генеративная .
Мегаспорогенез осуществляется в семязачатке . Из материнской клетки нуцеллуса в результате мейоза образуются 4 мегаспоры, из которых в результате остается только одна. Эта мегаспора сильно разрастается и оттесняет ткани нуцеллуса к интегументам, формируя зародышевый мешок . Ядро зародышевого мешка делится 3 раза митозом. После первого деления два дочерних ядра расходятся к разным полюсам: халазальному и микропилярному , и там делятся два раза. Таким образом, на каждом полюсе находится по четыре ядра. Три ядра у каждого полюса обособляются в отдельные клетки, а два оставшихся перемещаются в центр и сливаются, образуя вторичное диплоидное ядро. На микропилярном полюсе находятся две синергиды и одна более крупная клетка – яйцеклетка . На халазальном полюсе располагаются антиподы . Таким образом, зрелый зародышевыый мешок содержит 7 клеток.
Опыление заключается в переносе пыльцы с тычинок на рыльце пестика.
Оплодотворение . Пыльцевые зерна, тем или иным образом попавшие на рыльце, - прорастают. Прорастание пыльцы начинается с разбухания зерна и образования пыльцевой трубки из вегетативной клетки. Пыльцевая трубка прорывает оболочку в более тонком ее месте – так называемой апертуре. Кончик пыльцевой трубки выделяет специальные вещества, размягчающие ткани рыльца и столбика. По мере роста пыльцевой трубки в нее переходит ядро вегетативной клетки и генеративной, которая делится и образует два спермия. Через микропиле семязачатка пыльцевая трубка проникает в зародышевый мешок, там она разрывается, и ее содержимое изливается внутрь. Один из спермиев сливается с яйцеклеткой, образуя зиготу, которая затем дает начало зародышу семени. Второй спермий сливается с центральным ядром, что приводит к образованию триплоидного ядра, развивающегося затем в триплоидный эндосперм. Таким образом, эндосперм у покрытосеменных триплоидный и вторичный , т.к. образуется после оплодотворения.
Весь этот процесс получил название двойного оплодотворения . Он был впервые описан русским ученым С.Г.Навашиным. (1898 г.).
Антиподы и синергиды растворяются после оплодотворения, а интегументы преобразуются в семенную кожуру.
Апомиксис – развитие зародыша из неоплодотворенной клетки. Формы апомиксиса зависят от того, из каких частей семязачатка развивается зародыш. При партеногенезе (партенос- девственный) зародыш возникает из неоплодотворенной яйцеклетки. Если зародыш развивается из какой-либо другой клетки гаметофита (антиподы, синергиды), то такой процесс называется апогамия . В случае образования зародыша из клеток нуцеллуса, интегументов, не входящих в гаметофит, говорят об апоспории . Апомиксис распространен среди эволюционно развитых групп растений. За счет апомиксиса растения не зависят от опыляющих агентов
/>ДВОЙНОЕ ОПЛОДОТВОРЕНИЕ половой процесс у покрытосеменных растений, при к-ром оплодотворяются как яйцеклетка, так и центр. клетка зародышевого мешка. Д. о. открыл рус. учёный С. Г. Навашин в 1898 на 2 видах растений - лилии (Lilium martagon) и рябчике (Fritillaria orientalis) . В Д. о. участвуют оба спермия, привносимые в зародышевый мешок пыльцевой трубкой; ядро одного спермия сливается с ядром яйцеклетки, ядро второго - с полярными ядрами или со вторичным ядром зародышевого мешка. Из оплодотворённой яйцеклетки развивается зародыш, из центр. клетки - эндосперм. В зародышевых мешках с трёхклеточным лицевым аппаратом содержимое пыльцевой трубки обычно изливается в одну из синергид, которая при этом разрушается (в ней видны остатки ядра синер-гиды и вегетативного ядра пыльцевой трубки) ; вторая синергида впоследствии отмирает. Далее оба спермия вместе с изменённой цитоплазмой пыльцевой трубки перемещаются в щелевидный промежуток между яйцеклеткой и центр. клеткой. Затем спермин разобщаются: один из них проникает в яйцеклетку и вступает в контакт с её ядром, другой - проникает в центральную клетку, где контактирует со вторичным ядром или с одним, а иногда и с обоими полярными ядрами. Спермин теряют свою цитоплазму ещё в пыльцевой трубке или при проникновении в зародышевый мешок; иногда спермин в виде неизменённых клеток наблюдаются и в
/>
зародышевом мешке.
Двойное оплодотворение; 1 - у рябчика: один из спермиев (а) в контакте с ядром яйцеклетки, второй (б) - с одним из полярных ядер (второе полярное ядро не изображено); 2- у подсолнечника; а - пыльцевая трубка; 6 - синергиды (одна из них повреждена пыльцевой трубкой); в - яйцеклетка; г - спермий в контакте с ядром яйцеклетки; д - центральная клетка; е - второй спермий в контакте со вторичным ядром зародышевого мешка.
При Д. о. ядра зародышевого мешка находятся в интерфазе и обычно значительно крупнее ядер спермиев, форма и состояние к-рых могут вырьировать. У скерды и нек-рых др. сложноцветных ядра спермиев имеют вид двойной скрученной или извитой хроматиновой нити, у мн. растений они удлинённые, иногда извитые, б. или м. хроматизированные, не имеющие ядрышек; обычно спермии представляют собой округлые интерфазные ядра с ядрышками, иногда не отличающиеся по структуре от женских ядер.
По характеру объединения мужских и женских ядер предложено (Е. Н. Ге-расимова-Навашина) различать два типа Д. о.: премитотическое -ядро спермия погружается в женское ядро, хромосомы его деспирализуются; объединение хромосомных наборов обоих ядер происходит в интерфазе (в зиготе) ; постмитотическое - мужское и женское ядра, сохраняя свои оболочки, вступают в профазу, в конце к-рой начинается их объединение; интерфазные ядра, содержащие хромосомные наборы обоих ядер, образуются лишь после первого митотического деления зиготы. При Д. о. в яйцеклетке сливаются 2 гаплоидных ядра, поэтому ядро зиготы диплоид-но. Число хромосом в ядрах эндосперма зависит от числа полярных ядер в центр. клетке и от их плоидности; у большинства покрытосеменных 2 гаплоидных полярных ядра и эндосперм у них триплои-ден. Следствие Д. о. - Ксении - проявление доминантных признаков эндосперма отцовского растения в эндосперме гибридных семян. Если в зародышевый мешок проникает неск. пыльцевых трубок, спермий первой из них участвуют в Д. о., спермии остальных - дегенерируют. Случаи диспермии, т. е. оплодотворения яйцеклетки двумя спермиями, очень редки.
Рядом точных эмбриологических исследований на различных представителях покрытосеменных (лилейных, лютиковых, сложноцветных) С. Г. Навашин убедительно показал, что эндосперм, как и зародыш, является продуктом полового процесса. Это необычное, свойственное лишь покрытосеменным растениям, явление он назвал двойным оплодотворением. О своем открытии он сообщил в августе 1898 г. на проходившем в Киеве Х съезде русских естествоиспытателей и врачей, а в ноябре того же года опубликовал на эту тему небольшую статью в «Известиях Петербургской Академии Наук».
Мысль о существовании двойного оплодотворения возникла у С. Г. Навашина еще в 1895 г. в процессе работы над изучением халазогамии у грецкого ореха. Окончательное же подтверждение и оформление в стройную теорию эта мысль получила при исследовании оплодотворения у лилии.
Позднее С. Г. Навашин описал двойное оплодотворение и у других цветковых растений, систематически далеко отстоящих друг от друга - у представителей лютиковых, сложноцветных, орехоцветных, доказав тем самым общность этого явления для всех покрытосеменных.
Двойное оплодотворение - отличительный признак, отделяющий покрытосеменных от голосеменных.
Открытие С. Г. Навашиным двойного оплодотворения у покрытосеменных сыграло важную роль в науке.
Явление двойного оплодотворения внесло ясность не только в вопрос происхождения эндосперма, но и разъяснило загадочность такого явления, как ксения у кукурузы. Работы Навашина по оплодотворению покрытосеменных растений были встречены ботаниками всего мира с огромным интересом.
Дискуссии о сущности двойного оплодотворения продолжались и в начале ХХ в. В ходе этих дискуссий, а главное, благодаря новым исследованиям, выводы Навашина получили полное подтверждение и дальнейшее развитие.
половой процесс у покрытосеменных растений, при котором оплодотворяются как яйцеклетка, так и центральная клетка зародышевого мешка. Д. о. открыл русский учёный С.
Г. Навашин в 1898 на 2 видах растений ≈ лилии (Lilium martagon) и рябчике (Fritillaria orientalis). В Д. о. участвуют оба спермия, привносимые в зародышевый мешок пыльцевой трубкой; ядро одного спермия сливается с ядром яйцеклетки, ядро второго ≈ с полярными ядрами или со вторичным ядром зародышевого мешка. Из оплодотворённой яйцеклетки развивается зародыш, из центральной клетки ≈ эндосперм. В зародышевых мешках с трёхклеточным яйцевым аппаратом содержимое пыльцевой трубки обычно изливается в одну из синергид, которая при этом разрушается (в ней видны остатки ядра синергиды и вегетативного ядра пыльцевой трубки); вторая синергида впоследствии отмирает. Далее оба спермия вместе с измененной цитоплазмой пыльцевой трубки перемещаются в щелевидный промежуток между яйцеклеткой и центральной клеткой. Затем спермии разобщаются: один из них проникает в яйцеклетку и вступает в контакт с её ядром, другой ≈ проникает в центральную клетку, где контактирует со вторичным ядром или с одним, а иногда и с обоими полярными ядрами. Спермии теряют свою цитоплазму ещё в пыльцевой трубке или при проникновении в зародышевый мешок; иногда спермии в виде неизмененных клеток наблюдаются и в зародышевом мешке.
При Д. о. ядра зародышевого мешка находятся в интерфазе и обычно значительно крупнее ядер спермиев, форма и состояние которых могут вырьировать. У скерды и некоторых др. сложноцветных ядра спермиев имеют вид двойной скрученной или извитой хроматиновой нити, у многих растений они удлинённые, иногда извитые, более или менее хроматизированные, не имеющие ядрышек; обычно спермии представляют собой округлые интерфазные ядра с ядрышками, иногда не отличающиеся по структуре от женских ядер.
По характеру объединения мужских и женских ядер предложено (Е. Н. Герасимова-Навашина) различать два типа Д. о.: премитотическое ≈ ядро спермия погружается в женское ядро, хромосомы его деспирализуются; объединение хромосомных наборов обоих ядер происходит в интерфазе (в зиготе); постмитотическое ≈ мужское и женское ядра, сохраняя свои оболочки, вступают в профазу, в конце которой начинается их объединение; интерфазные ядра, содержащие хромосомные наборы обоих ядер, образуются лишь после первого митотического деления зиготы. При Д. о. в яйцеклетке сливаются 2 гаплоидных ядра, поэтому ядро зиготы диплоидно. Число хромосом в ядрах эндосперма зависит от числа полярных ядер в центральной клетке и от их плоидности; у большинства покрытосеменных 2 гаплоидных полярных ядра и эндосперм у них триплоиден. Следствие Д. о. ≈ ксении ≈ проявление доминантных признаков эндосперма отцовского растения в эндосперме гибридных семян. Если в зародышевый мешок проникает несколько пыльцевых трубок, спермий первой из них участвуют в Д. о., спермий остальных ≈ дегенерируют. Случаи диспермии, т. е. оплодотворения яйцеклетки двумя спермиями, очень редки.
Лит.: Навашин С. Г., Избр. труды, т. 1, М.≈ Л., 1951; Магешвар и П., Эмбриология покрытосеменных, пер. с англ., М., 1954; Поддубная Арнольди В. А., Общая эмбриология покрытосеменных растений, М., 1964; Steffen К., Fertilisation, в кн.: Maheshwari P. (ed.). Recent advances in the embryology of angiosperms, Delhi, 1963.
И. Д. Романов.
Двойное оплодотворение
Лекция Опыление
Для образования зародыша должны произойти опыление и оплодотворение.
Опыление — процесс переноса пыльцы с тычинки на рыльце пестика. Опыление впервые появляется у голосеменных растений, но наибольшего совершенства достигает у покрытосеменных.
Различают два типа опыления: самоопыление и перекрестное опыление. При самоопылении на пестик цветка попадает пыльца того же растения; если перенос пыльцы осуществляется между цветками разных особей, то происходит перекрестное опыление.
Считают, что оно свойственно 90% растений. Перекрестное опыление обусловливает высокий уровень гетерозиготности популяций. Это создает большие возможности для естественного отбора. Строгое самоопыление встречается относительно редко (например, у гороха) и может вести к расщеплению вида на ряд чистых линий, т.е. делает популяции гомозиготными.
Для эволюционного процесса считается оптимальным сочетание самоопыления и перекрестного опыления, что чаще и происходит в природе. Одна из форм, ограничивающих самоопыление, — двудомность, т.е. на одних растениях развиваются только мужские (тычиночные) цветки, а на других — женские (пестичные), имеющие только гинецей. Однодомные растения имеют цветки, содержащие и андроцей, и гинецей. Другая форма, ограничивающая самоопыление, — полная физиологическая несовместимость. Она выражается в подавлении при самоопылении прорастания пыльцы на рыльце пестика той же особи.
Выделяют два типа перекрестного опыления: биотическое и абиотическое. Биотическое опыление осуществляется животными. Энтомофилия — опыление насекомыми; орнитофилия — опыление птицами (колибри). Абиотическое опыление происходит с помощью неживых факторов внешней среды: ветром —анемофилия; водой у водных растений — гидрофилия.
Двойное оплодотворение
Пыльца, попав на рыльце пестика, начинает прорастать. Из вегетативной клетки образуется пыльцевая трубка, а из генеративной — два спермия. Пыльцевая трубка проникает в зародышевый мешок и, достигнув яйцеклетки, лопается, что обеспечивает проникновение в него спермиев. Один спермий копулирует с яйцеклеткой, образуя зиготу, дающую начало зародышу. Второй спермий сливается с вторичным диплоидным ядром, расположенным в центре зародышевого мешка, что приводит к образованию триплоидного ядра. В результате формируется триплоидная клетка, развивающаяся в специальную питательную ткань — эндосперм (э) (от греч. endon — внутри, sperma — семя). Таким образом происходитдвойное оплодотворение, свойственное только покрытосеменным растениям. Впервые его описал в 1898 г. Выдающийся русский цитологи эмбриолог С.Г. Навашин. Прочие клетки зародышевого мешка — антиподы и синергиды — разрушаются.
Биологический смысл двойного оплодотворения состоит в том, что триплоидный эндосперм развивается лишь в случае оплодотворения, чем достигается существенная экономия энергетических и пластических ресурсов, в отличие от голосеменных растений, у которых образование эндосперма не связано с оплодотворением.
У покрытосеменных растений эндосперм называют вторичным, или белком. Только у покрытосеменных растений зародыш (спорофаза) начинает свое развитие самостоятельно за счет триплоидной фазы. У всех предшествующих групп (голосеменных и др.) зародыш развивается за счет гаметофазы.
Таким образом, из составных частей цветка образуются:
Из оплодотворенной яйцеклетки — зародыш (2n);
Диплоидного ядра — эндосперм (3п);
Интегументов семязачатка — семенная кожура семени (2n);
Нуцеллуса — перисперм семени (2п);
Стенок завязи и часто с участием других элементов цветка (чашечки, цветоложа) — стенка плода (перикарпий).
Перикарпий состоит из трех слоев: наружного — экзокарпия, среднего — мезокарпия и внутреннего —эндокарпия.
У многих цветковых (около 10% видов) в процессе эволюции половое размножение замещается различными формами бесполого. Из них наиболее известен апомиксис, при котором семена у апомиктичных растений образуются без оплодотворения. При этом не происходит обмена генетической информацией, поэтому все апомиктические особи имеют одинаковую генетическую и соматическую конституцию. Наглядный пример растения с апомиктическим образованием семян по типу партеногенеза — одуванчик, отличающийся своей высокой жизнеспособностью. Нередко, особенно у многолетних корневищных растений, преобладает вегетативное размножение, а семенное подавлено.
СОЦВЕТИЯ
Соцветием называют побег или систему специализированных побегов, несущих цветки. Соцветия свойственны большинству цветковых растений. Они имеют главную ось (ось соцветия) и боковые оси. Боковые оси могут ветвиться или быть неразветвленными и нести цветки. На осях соцветий есть узлы и междоузлия. В узлах осей первого порядка расположены прицветники, а в узлах осей второго порядка — прицветнички.
Биологический смысл возникновения соцветия заключается в возрастающей вероятности опыления цветков как у энтомофильных, так и анемофильных растений. Несомненно, соцветия более заметны среди зелени листьев, нежели единичные цветки, и насекомое за единицу времени посетит гораздо больше цветков, если они собраны в соцветия. Обычно соцветия группируются близ верхней части растения, на концах ветвей, но иногда, особенно у тропических деревьев, возникают на стволах и толстых ветвях. Такое явление известно под названием каулифлории (от греч. kaulis — стебель и лат. flos — цветок). В качестве примера можно привести шоколадное дерево (Theobroma cacao). Считают, что в условиях тропического леса каулифлория делает цветки более доступными для насекомых-опылителей. Наличие соцветий предохраняет растение от бесплодия в случае гибели одиночного цветка.
В зависимости от степени разветвления осей различают простые и сложные соцветия. У простых соцветий на главной оси расположены одиночные цветки (черемуха, подорожник, подсолнечник); у сложных соцветий — не одиночные цветки, а боковые оси (донник лекарственный, сирень, пшеница, мятлик). У одних растений апикальные меристемы расходуются на формирование верхушечного цветка, и в этом случае соцветия относят к закрытым (симподиальным), или неопределенным. У закрытых соцветий верхушечные цветки обычно раскрываются раньше нижележащих боковых, и поэтому их называют верхоцветными. У других растений апикальные меристемы остаются в вегетативном состоянии, и такие соцветия называют открытыми (моноподиальными), или неопределенными. У открытых соцветий цветки распускаются последовательно снизу вверх, поэтому их называют бокоцветными.
Классификация соцветий достаточно сложна, но в справочниках и определителях учитывают два признака: характер ветвления и способ нарастания.
ПОСМОТРЕТЬ ЕЩЕ:
Цветковые (покрытосеменные) растения относятся к семенным растениям (наряду с голосеменными) и, следовательно, половое размножение у них осуществляется с помощью семян. При этом только у цветковых растений при половом размножении наблюдается такое явление как двойное оплодотворение. Оно было открыто в 1898 г. ученым С. Навашиным.
Суть двойного оплодотворения заключается в том, что у цветковых растений в оплодотворении участвуют два спермия. Один из них оплодотворяет яйцеклетку, в результате чего образуется зигота. Второй спермий оплодотворяет так называемую центральную клетку, из которой развивается запасающая ткань (эндосперм). При этом в зиготе восстанавливается двойной набор хромосом, а в будущем эндосперме - тройной (что уникально). Ниже процесс двойного оплодотворения у цветковых растений описан более подробно.
В тычинках, в их пыльцевых мешках, созревают пыльцевые зерна. Каждое пыльцевое зерно содержит две клетки: вегетативную и генеративную.
В завязи пестика развивается семязачаток (один, несколько или множество в зависимости от вида растения). Внутри семязачатка в результате деления образуются восемь клеток, содержащих одинарный набор хромосом (гаметофит). Две из этих клеток сливаются и образуется центральная клетка. Другая одна из этих клеток становится яйцеклеткой.
Когда пыльцевое зерно попадает на рыльце пестика, вегетативная клетка зерна и образует пыльцевую трубку, которая прорастает через ткани пестика и проникает в семязачаток.
Для этого в семезачатке есть специальное отверстие - пыльцевход.
Генеративная клетка пыльцевого зерна делится и образует два спермия. По пыльцевой трубке они проникают в семязачаток. Одни спермий оплодотворяет яйцеклетку, образуется зигота, содержащая двойной набор хромосом. Второй спермий сливается с центральной клеткой, в результате получается клетка с тройным набором хромосом.
В результате многочисленных делений зигота развивается в зародыш нового растения. В результате деления центральной клетки формируется эндосперм (питательная ткань для зародыша). Стенки семязачатка становятся семенной кожурой. Таким образом, семязачаток становится семенем.
Завязь пестика преобразуется в плод. Иногда в образовании плода участвует не только завязь, но и другие части цветка. Плод является своеобразным приспособлением цветковых растений к распространению семян. Разнообразие возможных способов распространения (с помощью животных, ветра, воды, саморазбрасыванием) породило огромное разнообразие плодов покрытосеменных растений.
Оплодотворение - это процесс слияния двух клеток, в результате чего происходит образование новой клетки, дающей начало другому организму этого же рода или вида. Что такое у цветковыхрастений и как оно происходит, читайте в данной статье.
Сущность оплодотворения
Оно происходит в результате слияния двух клеток, женской и мужской, и возникновения диплоидной зиготы. В каждой паре хромосом присутствует одна отцовская и одна материнская клетка. Сущность процесса оплодотворения заключается в том, чтобы восстановить и объединить наследственный материал родителей. Их потомство будет более жизнеспособным, так как соединит в себе самые полезные качества от отца и матери.
Оплодотворение - что такое?
Это процесс побуждения яйца к развитию в результате объединения ядер. Оплодотворение - что такое? Это необратимый процесс, который происходит в результате слияния разнополых гамет и объединения их ядер. не подвергается этой процедуре второй раз.
Но существуют растения, которые воспроизводят новое поколение только при помощи женской гаметы без оплодотворения. Такое размножение называется девственным. Примечательно, что эти два способа размножения у одного вида растений могут чередоваться.
Двойное оплодотворение цветковых растений
Обоих начал называются гаметами. Причем женскими являются яйцеклетки, а мужскими - спермии, которые у растений семенных неподвижные, а у споровых - подвижные. Оплодотворение - что такое? Это появление особой клетки - зиготы, содержащей наследственные признаки спермия и яйцеклетки.
Обладают сложным оплодотворением, которое называется двойным, поскольку, кроме яйцеклетки, оплодотворяется еще одна особая клетка. Формирование спермий происходит в пылинках пыльцы, а их созревание осуществляется в тычинках, точнее в их пыльниках. Местом образования яйцеклеток являются семязачатки, расположенные в завязи пестика. Когда яйцеклетка оплодотворится спермием, из семязачатка начинают развиваться семена.
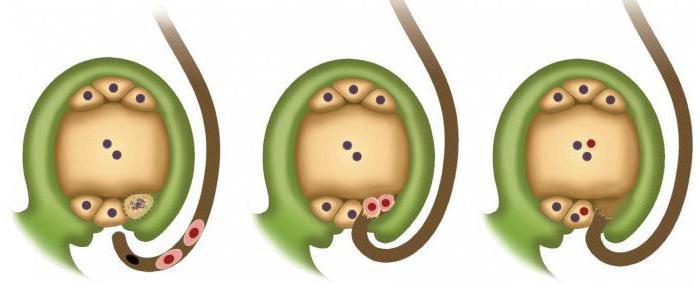
Чтобы оплодотворение у цветковых произошло, сначала нужно опылить растение, то есть на рыльце пестика должны попасть пылинки пыльцы. Оказавшись на рыльце, они начинают прорастать внутрь завязи, в результате чего образуется пыльцевая трубка. Одновременно с этим в пылинке происходит образование двух спермиев. Они не стоят на месте, а начинают продвигаться к пыльцевой трубке, которая проникает в семязачаток. Здесь в результате деления и удлинения одной клетки происходит образование зародышевого мешка.
Он нужен для расположения в нем яйцеклетки и еще одной клетки, в которой сосредоточен двойной набор наследственной информации. После этого происходит прорастание пыльцевой трубки в зародышевый мешок и слияние одного спермия с яйцеклеткой, в результате которого образуется зигота, а другого - с клеткой особой. Развитие зародыша происходит из зиготы. Второе слияние образует питательную ткань, или эндосперм, необходимый для питания зародыша в период роста.
Что нужно для существования каждого вида растений?
- Прежде всего необходимо восстановить диплоидный набор хромосом, а в его пределах - их парность.
- Обеспечить материальную непрерывность между поколениями, следующими чередой.
- Объединить в одном виде или роде наследственные свойства двух родителей.

Все это осуществляется на генетическом уровне. Для того чтобы оплодотворение осуществилось, созревание материнских и отцовских гамет должно произойти одновременно.
Оплодотворение у покрытосеменных растений
Этот процесс впервые охарактеризовал немецкий ученый Страсбургер во второй половине девятнадцатого века. Оплодотворение покрытосеменных растений происходит в результате слияния двух ядер разных гамет: с мужским и женским началом. Их цитоплазма не участвует в оплодотворении. Собственно оплодотворение происходит тогда, когда спермий сливается с ядром яйцеклетки.
Местом возникновения спермиев является пыльцевое зерно или пыльцевая труба. Зерно начинает прорастать после того, как попадает на рыльце. Время начала этого процесса у каждого растения разное, как и время оплодотворения. Например, пыльцевые зерна свеклы прорастают через два часа, а кукурузы - моментально. Первый признак прорастания зерна - его увеличение в объеме. Обычно одно пыльцевое зерно образует одну трубку. Но некоторые растения не подчиняются этому правилу и образуют несколько трубок, из которых только одна достигает своего развития.
Пыльцевая трубка с передвигающимися по ней спермиями растет и в конце концов разрывается. Все ее содержимое оказывается внутри зародышевого мешка. Один из проникших сюда спермиев внедряется в яйцеклетку и сливается с ее гаплоидным ядром. Оплодотворение - что такое? Это слияние двух ядер: спермия и яйцеклетки. Оплодотворенная яйцеклетка начинает делиться, получаются две новые клетки. Они делятся на четыре и так далее. Таким образом, происходит многократное деление, в результате которого развивается зародыш растения.

Покрытосеменные растения после процесса оплодотворения обладают способностью развивать дополнительный орган, который называется эндоспермом. Это не что иное, как питательная среда зародыша. При слиянии второго спермия и диплоидного ядра происходит образование определенного набора хромосом, из которых два - материнского происхождения, и один - отцовского. Таким образом, двойное оплодотворение организмов растительного происхождения осуществляется тогда, когда один спермий сливается с яйцеклеткой, а другой - с ядром клетки, расположенной в центре.
Отличительные черты покрытосеменных растений
- Большая приспособленность к произрастанию в разных условиях.
- Двойное оплодотворение, позволяющее иметь запас веществ, необходимых для нормального прорастания семян.
- Наличие триплоидного эндосперма.
- Образование семяпочек внутри завязи, при котором стенки пестика защищают их от повреждений.
- Развитие плода покрытосеменных растений из завязи.
- Нахождение семени внутри плода, стенки которого являются его защитой.
- Наличие цветка дает возможность насекомым.
Благодаря перечисленным признакам занимают господствующее положение в мире.
Особенность оплодотворения покрытосеменных растений
Она вытекает из того, что эти растения имеют двойное оплодотворение. Уникальная особенность представлена явлением, называемым ксениями. Его смысл заключается в том, что пыльца напрямую влияет на свойства и признаки эндосперма. Для примера возьмем кукурузу.

Она бывает с желтыми и белыми семенами. Их цвет зависит от оттенка эндосперма. При опылении женских цветков белозерной кукурузы пыльцой желтозерного сорта ее окраска все равно будет желтой, хотя развитие эндосперма происходит на растении с белыми зернами.
Какую роль играют цветковые растения?
Эти растения насчитывают 13 000 родов и 250 000 видов. Они получили широкое распространение по всему миру. Цветковые растения - это ключевые компоненты биосферы, производящие органические вещества, связывающие углекислоту и выделяющие кислород. Пастбищные цепи питания начинаются именно с них. Многие разновидности цветковых растений человек использует в пищу. Из них строят жилища и изготавливают различные хозяйственные материалы.

Не обходится без них и медицина. Отдельные виды покрытосеменных растений являются господствующими на планете, им отводится решающая роль в формировании покрова растительности и создании основной части наземной фитомассы. В конечном итоге именно этими растениями определяется возможность самого существования человека на земле как вида биологического.
Двойное оплодотворение у растений имеет большое биологическое значение. Оно было открыто Навашиным в 1898 г. Далее рассмотрим подробнее, как происходит двойное оплодотворение у растений.
Биологическое значение
Процесс двойного оплодотворения способствует активному развитию питательной ткани. В связи с этим семяпочка не запасает вещества впрок. Это, в свою очередь, объясняет ее быстрое развитие.
Схема двойного оплодотворения
Коротко явление можно описать следующим образом. Двойное оплодотворение у покрытосеменных растений состоит в проникновении в завязь двух спермиев. Один сливается с яйцеклеткой. Это способствует началу развития диплоидного зародыша. Второй спермий соединяется с центральной клеткой. В результате формируется триплоидный элемент. Из этой клетки появляется эндосперм. Он является питательным материалом для развивающегося зародыша.
Развитие пыльцевой трубки
Двойное оплодотворение у покрытосеменных начинается после образования гаплоидного сильно редуцированного поколения. Оно представлено гаметофитами. Двойное оплодотворение цветковых растений способствует прорастанию пыльцы. Оно начинается с разбухания зерна и последующего формирования пыльцевой трубки. Она прорывает спородерму в наиболее тонком ее участке. Называется он апертура. С кончика пыльцевой трубки выделяются специфические вещества. Они размягчают ткани столбика и рыльца. За счет этого в них входит пыльцевая трубка. По мере ее развития и роста, в нее переходят оба спермия и ядро от вегетативной клетки. В подавляющем большинстве случаев проникновение пыльцевой трубки в нуцеллус (мегаспорангий) происходит посредством микропиле семязачатка. Крайне редко это осуществляется другим способом. После проникновения в зародышевый мешок происходит разрыв пыльцевой трубки. В результате все ее содержимое изливается вовнутрь. Двойное оплодотворение цветковых растений продолжается формированием диплоидной зиготы. Этому способствует первый спермий. Второй элемент соединяется с вторичным ядром, которое расположено в центральной части зародышевого мешка. Образованное триплоидное ядро впоследствии трансформируется в эндосперм.

Формирование клеток: общие сведения
Процесс двойного оплодотворения цветковых растений осуществляется особыми половыми клетками. Их формирование происходит в два этапа. Первая стадия называется спорогенез, вторая - гематогенез. В случае образования мужских клеток эти этапы именуются микроспорогенез и микрогематогенез. При образовании женских половых элементов приставка меняется на "мега" (или "макро"). Спорогенез основывается на мейозе. Это процесс формирования гаплоидных элементов. Мейозу, так же как и у представителей фауны, предшествует размножение клеток посредством митотических делений.
Образование спермиев
Первичное формирование мужских половых элементов осуществляется в особой ткани пыльника. Она называется археспориальной. В ней в результате митозов происходит формирование многочисленных эелементов - материнских клеток пыльцы. Они и вступают затем в мейоз. Вследствие двух мейотических делений образуется 4 гаплоидные микроспоры. Некоторое время они лежат рядом, формируя тетрады. После этого происходит их распад на пыльцевые зерна - отдельные микроспоры. Каждый из образованных элементов начинает покрываться двумя оболочками: внешней (экзина) и внутренней (интина). Затем начинается следующий этап - микрогаметогенез. Он, в свою очередь, состоит из двух митотических последовательных делений. После первого формируется две клетки: генеративная и вегетативная. Впоследствии первая проходит еще одно деление. В результате образуется две мужские клетки - спермии.

Макроспорогенез и мегаспорогенез
В тканях семяпочки начинает обособляться один или несколько археспориальных элементов. Они начинают усиленно расти. Вследствие такой активности они становятся значительно крупнее остальных клеток, окружающих их в семяпочке. Каждый археспориальный элемент один, два или более раз подвергается делению митозом. В некоторых случаях клетка может сразу трансформироваться в материнскую. Внутри нее происходит мейоз. В результате него формируется 4 гаплоидные клетки. Как правило, самая крупная из них начинает развиваться, превращаясь в зародышевый мешок. Три оставшиеся постепенно дегенерируют. На данном этапе макроспорогенез завершается, начинается макрогематогенез. В ходе него происходят митотические деления (у большей части покрытосеменных их три). Цитокинез не сопровождает митозы. В результате трех делений формируется зародышевый мешок с восемью ядрами. Они впоследствии обосабливаются в самостоятельные клетки. Эти элементы распределяются определенным образом по зародышевому мешку. Одна из обособленных клеток, которая, собственно, является яйцеклеткой, совместно с двумя другими - синергидами, занимает место у микропиле, в которое осуществляется проникновение спермиев. В этом процессе синергиды исполняют очень значимую роль. В них содержатся ферменты, которые способствуют растворению оболочек на пыльцевых трубках. В противоположной стороне зародышевого мешка располагаются другие три клетки. Они именуются антиподами. С помощью этих элементов происходит передача из семяпочки питательных веществ в зародышевый мешок. Оставшиеся две клетки располагаются в центральной части. Зачастую они сливаются. В результате их соединения формируется диплоидная центральная клетка. После того как произойдет двойное оплодотворение, и в завязь проникнут спермии, один из них, как выше сказано было, сольется с яйцеклеткой.

Особенности пыльцевой трубки
Двойное оплодотворение сопровождается взаимодействием ее с тканями спорофита. Оно достаточно специфично. Этот процесс регулируется активностью химических соединений. Установлено, что если пыльцу промыть в дистиллированной воде, она потеряет способность к прорастанию. Если же полученный раствор сконцентрировать, а затем ее обработать, то она снова станет полноценной. Развитие пыльцевой трубки после прорастания контролируют ткани пестика. К примеру, у хлопчатника ее рост до яйцеклетки занимает порядка 12-18-ти часов. Однако уже спустя 6 часов вполне можно определить, к какой именно семяпочке будет направляться пыльцевая трубка. Это понятно потому, что в ней начинается разрушение синергиды. В настоящее время не установлено, как растение может направить развитие трубки в нужном направлении и каким образом о приближении узнает синергида.

"Запрет" на самоопыление
Он достаточно часто наблюдается у цветковых растений. Это явление имеет свои особенности. "Запрет" на самоопыление проявляется в том, что спорофит "идентифицирует" собственного мужского гематофита и не допускает его к участию в оплодотворении. При этом в ряде случаев на рыльце пестика не происходит прорастания собственной пыльцы. Однако, как правило, рост трубки все-таки начинается, но впоследствии приостанавливается. В результате пыльца не достигает яйцеклетки и, как следствие, двойное оплодотворение не происходит. Еще Дарвиным было отмечено это явление. Так, он обнаружил у первоцвета весеннего цветки двух форм. Одни из них были длинностолбиковыми с короткими тычинками. Другие же - короткостолбиковыми. В них тычиночные нити были длинные. Короткостолбиковые растения отличаются крупной пыльцой (вдвое больше, чем у других). При этом клетки в сосочках рыльца - мелкие. Указанные признаки контролирует группа из тесно сплетенных генов.

Рецепторы
Двойное оплодотворение эффективно, когда пыльца переносится от одной формы к другой. За распознавание собственных элементов отвечают особые молекулы-рецепторы. Они представляют собой сложные соединения углеводов с белками. Установлено, что формы дикой капусты, не вырабатывающие в тканях рыльца эти молекулы-рецепторы, способны самоопыляться. Для нормальных растений характерно появление углеводно-белковых соединений за день до раскрытия цветка. Если открыть бутон и обработать его собственной пыльцой за двое суток до его распускания, то двойное оплодотворение произойдет. Если это сделать за день до открытия, то его не будет.
Аллели
Примечательно, что в ряде случаев "самонесовместимость" пыльцы в растениях устанавливается серией множественных элементов одного гена. Это явление похоже на несовместимость при пересадке ткани у животных. Такие аллели обозначают литерой S. Число в популяции этих элементов может достичь десятков или даже сотен. К примеру, если генотип растения, производящего яйцеклетки, - s1s2, а вырабатывающего пыльцу - s2s3, при перекрестном опылении прорастание будет отмечено только у 50% пылинок. Это будут те, которые несут аллель s3. Если элементов несколько десятков, то большая часть пыльцы прорастет нормально при перекрестном опылении, при этом самоопыление предотвращается полностью.

В заключение
В отличие от голосеменных, для которых характерно развитие достаточно мощного гаплоидного эндосперма вне зависимости от оплодотворения, у покрытосеменных ткань образуется только в этом единственном случае. Учитывая огромное количество поколений, таким образом достигается значительная экономия энергии. Повышение степени плоидности эндосперма, по всей видимости, способствует более скорому росту ткани в сравнении с диплоидными слоями спорофита.
>>Оплодотворение у цветковых растений
Получая из внешней среды все необходимое для жизни, цветковые растения растут, цветут и образуют плоды с семенами. Чтобы завязался плод и развились семена, должно произойти опыление , а после него - оплодотворение.
Оплодотворением называют слияние двух половых клеток - гамет. У цветковых растений мужские гаметы - спермии - очень мелкие. Женские гаметы - яйцеклетки - гораздо крупнее спермиев.
При опылении пылинки, или пыльцевые зерна, попадают на рыльца. Внешне пыльцевые зерна разных растений очень разнообразны, у многих растений они имеют форму маленьких шариков. Каждое пыльцевое зерно одето оболочкой, поверхность которой редко гладкая; чаще она неровная и покрыта шипиками, бородавочками, выростами в виде сеточки 9 3 . Это помогает пыльцевым зернам удерживаться на теле насекомого-опылителя и на рыльце.
На поверхности рыльца выделяется липкая, удерживающая пыльцу жидкость. Здесь пылинка прорастает в длинную, очень тонкую пыльцевую трубку. Пыльцевая трубка сначала растет между клетками рыльца, затем - столбика и наконец врастает в полость завязи.
В полости завязи находятся семязачатки (семяпочки). Число семязачатков в завязях разных растений различно. У пшеницы , ячменя, ржи, вишни завязь содержит только один семязачаток, V хлопчатника - несколько десятков а у мака их число достигает нескольких тысяч.
По мере роста пыльцевой трубки по ней из пыльцевого зерна перемещаются две Клетки , имеющие крупные ядра . Это спермин. Они все время находятся близ растущего кончика пыльцевой трубки 94 .
Семязачатки развиваются на внутренних сторонах стенок завязи и, как все части растения, состоят из клеток. Каждый семязачаток одет покровом, в котором на вершине семязачатка.
В этой ткани, состоящей из мелких клеток с тонкими оболочками, развивается группа относительно крупных клеток. Среди них ближе к пыльцевходу находится яйцеклетка. Кончик пыльцевой трубки врастает в семязачаток. Яйцеклетка сливается с одним из спермиев. Происходит оплодотворение.
Второй спермин сливается с самой крупной из группы клеток семязачатка. Таким образом, у цветковых растений при оплодотворении происходит два слияния: первый спермий сливается с яйцеклеткой, второй - с крупной центральной клеткой. Этот процесс открыл в 1898 г. русский ботаник, академик С. Г. Навашин и назвал его двойным оплодотворением.
1. Что такое опыление?
2. Что называют оплодотворением?
3. Где находятся семязачатки?
4. Сколько семязачатков бывает в завязях разных растений?
5. Где находится яйцеклетка?
6. Как происходит оплодотворение у цветковых растений?
Корчагина В. А., Биология: Растения, бактерии, грибы, лишайники: Учеб. для 6 кл. сред. шк. - 24-е изд. - М.: Просвещение, 2003. - 256 с.:ил.
Календарно-тематическое планирование по биологии, видео по биологии онлайн , Биология в школе скачать
Содержание урока конспект урока опорный каркас презентация урока акселеративные методы интерактивные технологии Практика задачи и упражнения самопроверка практикумы, тренинги, кейсы, квесты домашние задания дискуссионные вопросы риторические вопросы от учеников Иллюстрации аудио-, видеоклипы и мультимедиа фотографии, картинки графики, таблицы, схемы юмор, анекдоты, приколы, комиксы притчи, поговорки, кроссворды, цитаты Дополнения рефераты статьи фишки для любознательных шпаргалки учебники основные и дополнительные словарь терминов прочие Совершенствование учебников и уроков исправление ошибок в учебнике обновление фрагмента в учебнике элементы новаторства на уроке замена устаревших знаний новыми Только для учителей идеальные уроки календарный план на год методические рекомендации программы обсуждения Интегрированные урокиРастения - они такие загадочные. Ну всё у них не так, как у людей. Даже половые клетки они образуют не мейозом, а митозом, да еще и чередование поколений, да еще и оплодотворение двойное...
Впрочем, обо всём по порядку...
Вспомним строение цветка
У покрытосеменных растений процесс формирования половых клеток состоит из двух этапов: спорогенеза и гаметогенеза. Пыльца образуется в пыльниках тычинок. Микроспорогенез – процесс образования микроспор в микроспорангиях (гнезда пыльника), где в результате митозов возникают материнские клетки пыльцы, которые вступают в мейоз. После двух мейотических делений образуется 4 гаплоидные микроспоры– пыльцевые зерна. Пыльцевое зерно (пылинка) покрыто двумя оболочками. Затем внутри пыльцевого зерна происходит микрогаметогенез (процесс образования мужского гаметофита из микроспоры) – два последовательных митотических деления. В результате первого образуются вегетативная и генеративная клетки, а после второго деления из генеративной клетки образуются два спермия.
Макроспорогенез (процесс формирования мегаспор) происходит в семязачатках, которые расположены в завязи пестика (покрыты). В области микропиле начинает разрастаться одна клетка – мегаспороцит, или материнская клетка мегаспор. В ней происходит мейоз , и образуются 4 гаплоидные клетки. Одна из этих клеток развивается в зародышевый мешок, три остальных разрушаются. Далее начинается макрогаметогенез – формирование женского гаметофита. После трех митотических делений образуется восьмиядерный зародышевый мешок. Ядра в дальнейшем обособляются в самостоятельные клетки. Одна из этих клеток (яйцеклетка) с двумя клетками-синергидами располагается возле микропиле. Синергиды содержат ферменты, которые растворяют оболочку пыльцевой трубки и при необходимости могут заместить яйцеклетку. На противоположной части зародышевого мешка располагаются три клетки-антиподы (передатчики питательных веществ из семяпочки в зародышевый мешок). Две оставшиеся клетки сливаются и образуют крупную центральную диплоидную клетку. Зародышевый мешок имеет покровы.


Процессу оплодотворения предшествует процесс опыления. Попав на рыльце, пыльцевое зерно прорастает в пыльцевую трубку, в которой расположены два неподвижных спермия. Синергиды выделяют ферменты, разрушающие пыльцевую трубку, и ее содержимое изливается рядом с яйцеклеткой. Оплодотворение от воды не зависит. Двойное оплодотворение (С.Г. Навашин, 1898 г.): один спермий сливается с яйцеклеткой (образуется зигота), второй – с диплоидной центральной клеткой, образуя триплоидный эндосперм. Из зиготы далее развивается зародыш, покровы превращаются в семенную кожуру. Развивающееся семя защищено стенками завязи (околоплодник). На месте цветка созревает плод.